www.joiiup.com
低能量光療(LLLT)對腦部疾病的機制


憂鬱症和阿茲海默症(AD)患者逐年增加,已逐漸演變成全球流行病的程度,對大腦健康的關注已成為多數人的重要大事。從藥物到冥想,人們正在尋找改善大腦並使其保持一輩子健康的方法。而低能量光療法(Low-Level Laser/Light Therapy; LLLT)可能是你甚至你的醫生都不知道的一種改善你大腦疾病的有效療法。

LLLT使用於醫療已經半個多世紀,目前已在物理療法和康復療法中確立了完善的地位,現在光的治療用途擴大到包括中風、心肌梗塞、某些腦部疾病和頭部外傷(TBI)。並且有望用於治療其他大腦問題的研究正在進行中,例如憂鬱症、焦慮症、自閉症、成癮、帕金森氏症和阿茲海默症。
LLLT經過廣泛且深度的研究後,發現是安全有效、且不具有害副作用的治療方式。與使用於手術的高強度雷射不同,LLLT不會切割或燃燒人體組織,而且LLLT可以促進細胞發揮最佳的功能而達到疾病的治療。本文即針對已被研究證實的LLLT對大腦疾病治療的各種機制進行系統化整理。
細胞色素c氧化酶(cytochrome c oxidase ; CCO)
對LLLT作用機制最深入研究的重點就是細胞色素c氧化酶(cytochrome c oxidase ; CCO),它是線粒體呼吸鏈的第四個單元(如圖一),負責利用葡萄糖代謝所產生的電子將氧氣還原為水[1]。從理論上講,一氧化氮(NO)可能會抑制CCO酶的活性(尤其是在缺氧或受損的細胞中)。這種具抑制性的NO可以被CCO(包含兩個血紅素和兩個擁有不同吸收光譜的銅中心CuA和CuB)所吸收的光子從CCO解離[2]。這些吸收峰主要在紅色(600-700 nm)和近紅外線(760-940 nm)光譜區域。當NO被光離解釋放出時,線粒體膜電位會增加,使更多的氧氣被消耗、更多葡萄糖被代謝,而線粒體會產生更多的ATP。
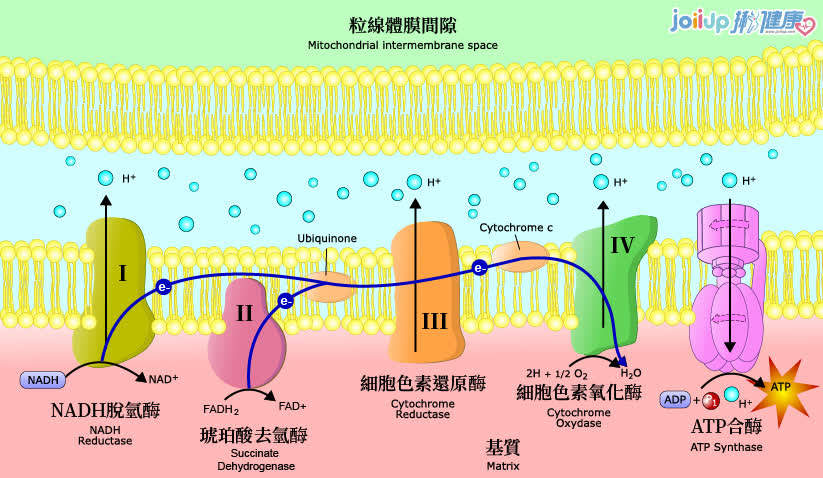
圖一:線粒體內的電子傳遞鏈(呼吸鏈)第四個單元就是細胞色素氧化酶
代謝功能改善是LLLT最容易被察覺的作用之一,而增加的細胞內ATP產生則是最受支持的作用機制之一[3]。此外,一些臨床前研究表明,由於各種腦部疾病,接受LLLT的實驗動物(小鼠或大鼠)的大腦ATP含量明顯增加了[4, 5]。普遍認為線粒體功能障礙、ATP供應不足和氧化壓力是幾乎所有形式的腦疾病的促成因素[6]。據研究報導,這涉及神經系統疾病,例如重度憂鬱症、腦外傷、帕金森氏病和AD [7-10]。
活性氧(Reactive oxygen species; ROS)、一氧化氮、血流
研究顯示當線粒體吸收LLLT期間傳遞的光子時,線粒體中產生的活性氧(ROS)會瞬間增加。ROS的爆發可能觸發線粒體信號傳導途徑,從而導致細胞中的細胞保護、抗氧化和抗凋亡(anti-apoptotic)作用[11]。通過光解離作用釋放的NO既可作為血管擴張劑,也可作為淋巴液的擴張劑。此外,NO也是有效的信號分子,可以激發許多有益的細胞途徑[12],圖二說明了這些機制。
LLLT光照後最容易測量的變化之一是腦血流量和氧合的變化。以1064 nm近紅外線來照射志願者的前臂[13],發現隨著時間的推移、光能量劑量的累積,LLLT引起照射部位的CCO濃度(Delta [CCO])和氧化血紅蛋白濃度(Delta [HbO])明顯增加。有人提出,由於LLLT導致的NO釋放是使腦血流量增加的原因[14]。 NO是主要的神經元信號分子,具有觸發血管舒張的能力。
為此,它首先刺激可溶性鳥苷酸環化酶(guanylate cyclase)形成環磷酸鳥苷(cyclic GMP; cGMP),然後cGMP激發蛋白激酶G,導致鈣離子的再攝取和鈣激發鉀通道的開放。隨後鈣離子濃度下降可防止肌球蛋白輕鏈激酶(myosin light-chain kinase)被磷酸化成肌球蛋白分子,進而使血管和淋巴管內壁的平滑肌細胞鬆弛[15]。然後,這種血管舒張促進了血液循環的改善,也改善了大腦氧合。而腦血流量(cerebral blood flow ; CBF)疾病、神經血管功能障礙和較低的大腦氧合水平已被認為是AD的重要特徵[16]。
光敏離子通道和鈣
除了CCO以外,還必須有其他類型的光感受器,因為某些研究明顯證明即使比上述紅光/近紅外線更長的波長也會產生有益的影響。據報導,諸如980 nm [17]、1064 nm雷射[18]和1072 nm LED [19]等波長,甚至寬帶IR光[20]都具有光生物調變類型的效應。
儘管尚未確定這些波長的光感受器,但目前主要的論點認為是位於熱敏感或光敏感離子通道中的水(或許是奈米結構水)。藉由光介導的鈣離子通道的打開可以用來解釋觀察到的細胞內鈣明顯變化,例如瞬間受器電位(transient receptor potential ; TRP)超家族的成員[21]。TRP描述了一個以瞬間受器電位陽離子通道 (transient receptor potential cation channel, subfamily V, member 1 ;TRPV1為典型的大離子通道家族,最近被確定為是辣椒素(辣椒中的活性成分)的生物受體[22]。TRP通道的生物學作用是多種多樣的,但是許多TRP通道涉及到熱感測和溫度調節[23]。
信號傳導介質和轉錄因子激發
大多數研究團隊認為,LLLT對大腦的有益作用可以藉由腦血流量的增加、更多的氧氣供應、氧氣消耗、ATP產生和線粒體活性改善來解釋[24, 25]。有許多研究表明,短暫的光照(尤其是遭受某種急性損傷或外傷的實驗動物)的效果可能會持續數天、數週甚至數月[26]。光的這種長效作用只能藉由激發導致長時間蛋白質表達變化的信號傳導途徑和轉錄因子來解釋。LLLT對刺激線粒體活性和血流的影響本身無法解釋光照的長期影響。
最近的綜合研究列出了不低於十四種不同的轉錄因子和信號傳導介質,據報導它們在光曝照後會被激發[1]。圖二說明了吸收光子的腦部神經元細胞內兩個最重要的分子感光體或生色團(細胞色素c氧化酶和熱控離子通道)。信號傳導途徑和轉錄因子的激發導致LLLT在大腦中的最終作用。
圖三說明了一些組織特定的機制,這些機制是由圖二中解釋的初始光子吸收效應所引起。緊接著可以發生各種流程效應,這些效應可以使相應範圍的腦部疾病受益。這些效應可分為短期刺激(ATP、血流、淋巴流、腦部氧合、水腫減少)。另一組流程效應則圍繞著神經保護(抗凋亡蛋白的上調、神經興奮毒性降低、抗氧化劑增加、發炎減少),其可以歸為“幫助大腦自我修復”的效應,包含神經營養蛋白(neurotrophins)、神經元生成(neurogenesis)和突觸生成(synaptogenesis)。

圖二:LLLT的分子和細胞內機制。 AP1(激發蛋白1)、 ATP(三磷酸腺苷)、 Ca2+(鈣離子)、cAMP(環狀單磷酸腺苷)、NF-kB(核因子κB)、NO(一氧化氮)、ROS(活性氧)、TRPV(瞬態感受器電位陽離子通道)。(圖片內容來源:M.R. Hamblin / BBA Clinical 6 (2016) 113–124,JoiiUp 重新製圖)

圖三:LLLT療程後發生的組織特異性過程,有益於一系列腦部疾病。腦源性神經營養因子(BDNF);神經生長因子(NGF);神經營養蛋白3 (neurotrophin 3;NT-3);超氧化物歧化酶(superoxide dismutase ;SOD)。(圖片內容來源:M.R. Hamblin / BBA Clinical 6 (2016) 113–124,JoiiUp 重新製圖)
神經保護
各種證據表明LLLT可以用於神經保護,他透過抑制糖原合酶激酶3β (Glycogen synthase kinase 3 beta ; GSK3β)的活性來保護細胞免受損傷,促進其存活和壽命以及逆轉凋亡信號傳導過程,防止細胞凋亡,凋亡是隨著生物體生長而發生的正常細胞死亡。據信GSK3β充當AKT和Bax凋亡基因之間的介體,Bax是一種在促凋亡刺激存在下易位至細胞核以觸發該過程開始的蛋白質。但是,當GSK3β受到抑制時,AKT與AKT之間的通信路徑被切斷。結果,無法表達Bax易位,因此被抑制[27]。
LLLT還以防止衰老的形式表現出神經保護作用[28]。已顯示它可以激發細胞外信號相關激酶(extracellular signal-related kinase ; ERK)/前叉箱蛋白M1(forkhead box protein M1; FOXM1)途徑。FOXM1蛋白調節細胞週期從G1到S期,並且透過激發ERK / FOXM1途徑,LLLT導致ERK向細胞核的轉運更大、FOXM1在核中的積累更大。反過來,這會導致p21蛋白的表達降低以及G1期的有絲分裂停滯,從而減慢了細胞衰老的進程。
LLLT也被證實可有效保護細胞免受毒素傷害[29]。在Eells團隊的研究裡[30]暴露於甲醇的囓齒動物模型中,用670 nm光照成功地使視網膜功能恢復並防止了組織損傷。這可能是由於以下事實:甲醇會生成有毒的代謝產物甲酸(CCO的抑制劑),而LLLT是CCO已知的刺激劑,LLLT的抗毒素作用可以追溯到其對CCO的刺激作用。
此外,LLLT已證明可用不同的方式影響處於不同健康狀態的細胞,本質上是以促進其存活所必需的方式修飾細胞。例如,在正常細胞中,CCO對光的吸收會導致基質金屬蛋白酶(Matrix Metalloproteinase; MMP) 高於基線並導致ROS產生短暫增加。然而,在存在氧化壓力,興奮性毒性或抑制電子傳輸而導致MMP較低的細胞中,光吸收會導致MMP朝正常濃度增加,並降低ROS產生[31]。同樣,健康細胞對LLLT的典型反應是細胞內鈣離子的短暫增加[32]。然而,在已經含有過量鈣離子的細胞中(一種稱為興奮性毒性的現象),LLLT會引發相反的反應,換言之,它會降低細胞鈣的過量濃度,從而促進細胞存活、降低氧化壓力並使MMP恢復至正常水準 [33]。
氧化壓力
當人體抵抗活性氧作用的能力與活性氧(ROS)產生發生不平衡的狀況時,就會發生氧化壓力,當活性氧過剩,它們會透過抗氧化劑而變得有害。許多研究顯示氧化壓力與各種神經系統疾病相關,例如嚴重的憂鬱症和顱腦外傷,更不用說心血管疾病和阿茲海默症[34-37]。但是,這種情況比起初步觀察要來的複雜得多,因為大量針對這些疾病的抗氧化劑治療臨床試驗都失敗了[38]。顯然,一定水準的氧化壓力對於人類的最佳運作功能是必需的,而透過補充抗氧化劑去除所有氧化壓力可能適得其反[39]。一篇重要的論文表明,服用抗氧化劑時,運動對健康的好處就會消失[40]。
Salehpour團隊 [41]表明,小鼠的睡眠剝奪(SD)會引起海馬的氧化壓力,進而導致記憶障礙。將具有NIR(810 nm)的LLLT經顱治療(每天一次,連續3天)遞送至頭部,小鼠在Barnes迷宮和What-Where-Which任務上表現更好,海馬中抗氧化酶濃度增加、氧化壓力生物標誌物降低。在研究LLLT對受創傷肌肉的影響時,證明其可有效調節細胞產生的細胞因子誘導型一氧化氮合酶(cytokine-inducible nitric oxide synthase ; iNOS)的量。這個很重要,因為過量的iNOS會導致NO的過量生成,這將引發ROS / RNS(過氧亞硝酸鹽)的生成增加,從而導致氧化壓力增加。具體而言,LLLT可以減少過氧亞硝酸鹽[42],同時仍保留NO合酶其他亞型的積極作用,例如內皮型一氧化氮合酶(endothelial nitric oxygen synthase ; eNOS),這是主要負責LLLT血管舒張作用的成份[43, 44] 。
LLLT也已顯示出刺激血管生成的增加,從而導致血流的進一步改善。正如Cury [45]所證明的,在780 nm和40J/cm2的LLLT觸發了蛋白HIF1α和血管內皮生長因子表達的增加,以及基質金屬蛋白酶2 (matrix metalloproteinase 2)活性的降低,這些都被發現能誘導血管生成。另外,在體外研究紅/近紅外線對紅細胞的影響時,發現近紅外線在保護紅細胞免於氧化方面非常有效[46],氧化是受如嚴重憂鬱症等疾病折磨的大腦中的常見現象[47]。廣泛的線粒體功能障礙、鋁和重金屬含量增加以及AD中發生的神經發炎會產生大量的氧化壓力。氧化壓力導致Aβ沉積,tau蛋白過度磷酸化以及隨後的突觸和神經元喪失[48]。
抗發炎效應
發炎是先天免疫系統對細菌和病毒等異物的防禦之一,在細胞層級上,它會在轉錄因子(transcription factor) NF-κB被激發時發生。雖然急性發炎是有助於人體的,但慢性發炎就會產生非常不利的影響。許多疾病,包括神經退行性疾病和情緒障礙,可以部分歸因於慢性發炎。LLLT幫助平息發炎的一種方法是抑制環加氧酶2(cyclo-oxygenase 2; COX-2)酶。 Lim研究團隊 [49]發現低功率的635 nm光照射能夠通過減少細胞內ROS引起COX-2抑制。目前,廣泛支持利用藥物來抑制COX-2,其中COX-2抑制劑在非類固醇類抗炎藥市場上佔有很大的份額[50],而基本上使用LLLT就可以實現相同的結果。
LLLT還可以調節細胞中游離的NF-κB水平。NF-κB可以被發現在與抑制蛋白IκB結合的細胞質中。促炎性刺激具有激發IκB激酶的能力,激酶是引起IκB降解的上游信號調節劑。IκB降解後,NF-κB可以自由轉運至細胞核,從而觸發促炎基因的表達。有證據表明,LLLT可以對NF-κB產生相反的影響,具體取決於研究的細胞類型及其激發狀態。研究發現在正常的成纖維細胞中,LLLT可以藉由刺激線粒體產生少量ROS來激發NF-κB [51]。然而,同一小組發現在樹突狀細胞(另一種巨噬細胞)中,該細胞已被激發為M1表型,LLLT可以減少促炎細胞因子[52]。同樣,Yamaura團隊發現在接受LLLT的活化類風濕性關節炎滑膜細胞中NF-κB濃度降低了[53]。
另外,LLLT具有調節細胞激素(cytokines)濃度的能力,這些蛋白是免疫系統的重要信號分子。研究顯示LLLT可調節促炎和抗炎細胞激素的濃度,儘管對於減少發炎,其調節腫瘤壞死因子(tumor necrosis factor ;TNF)和其他促炎細胞激素濃度的能力特別有用。應該注意的是,與身體其他部位的發炎相比,大腦內的發炎具有明顯的差異。實際上,“神經發炎(neuroinflammation)” 這個術語通常用於激發小膠質細胞(microglia)。小膠質細胞在中樞神經系統中起免疫防禦系統的作用[54]。小膠質細胞不斷清除中樞神經系統的斑塊,受損的神經元和突觸以及傳染原。
小膠質細胞可以呈現多種表型,並保留改變其功能以維持組織穩態的能力。小膠質細胞可以藉由LPS或IFN-γ激發成表達促炎性細胞因子並能殺死微生物細胞的M1表型。另一方面,小膠質細胞可以被IL-4 / IL-13激發為M2型,以吞噬碎片、消炎和修復組織。越來越多的證據表明,代謝重新編程在先天性發炎反應的調節中起著作用[55]。既然有大量證據表明LLLT可以激發線粒體代謝,使其氧化為磷酸化(oxidative phosphorylation),並且遠離有氧糖酵解,這就是LLLT可能將小膠質細胞表型(microglial phenotype)從M1變為M2的合理原因[56]。這種轉變的結果是LLLT誘導的M2可以代替M1,該M1無法處理AD中的β-澱粉樣斑塊等物質,因此會產生ROS和炎性細胞因子。而LLLT誘導的M2可以清除斑塊,發揮抗炎和抗氧化作用,並促進組織癒合[57]。
神經生成
多年來,人們一直認為成年大腦無能力生長新的腦細胞。儘管已經認識到,胚胎、幼小動物和兒童大腦的成長和發育必須能夠由神經幹細胞(neural stem cells ; NSC)和神經祖細胞(neuroprogenitor cells)介導的神經發生,儘管這一過程在成年期就已經停止了。我們認識的轉折點是發現了成人的神經生成,並鑑定了在體外和體內均可作為NSC發揮作用,其能產生新的神經元、神經膠質細胞[58]。
Oron等人的一項研究首次報導了通過LLLT刺激腦部神經所發生的神經生成。在2006年,他在大鼠中誘發了中風,並用LLLT治療了它們。當用LLLT治療時,在半球同側腦室下區域新形成的神經元細胞和遷移細胞的數量顯著增加[59]。Xuan團隊研究也發現了類似的結果,他們使用LLLT治療了患有頭部外傷的小鼠[60]。
促進突觸生成(Synaptogenesis)
迄今為止,LLLT對大腦最顯著且潛在的影響之一是其促進突觸生成的能力,也稱為神經可塑性。這個過程至關重要,因為可以將包括頭部外傷、中風、神經退行性疾病和情緒障礙在內的許多大腦狀況部分或全部追溯到大腦某些區域的神經元連接不良或異常。如果LLLT具有通過促進神經組織或重組來抵抗這些作用的能力,那麼它可能被證明是治療這類型腦部疾病的一種新方法。
LLLT促進神經元連接的一種方式可能是藉由上調腦源性神經營養因子(brain derived neurotrophic factor ; BDNF)。BDNF是神經系統中發現的一種蛋白質,有助於維持現有的神經元並促進新神經元和新突觸的生長。據信調節樹突結構可以促進改善的突觸傳遞[61]。LLLT已顯示可通過ERK / CREB (cAMP-response element binding protein; 環磷腺苷效應元件結合蛋白) 途徑減慢BDNF的衰減,從而積極影響樹突形態發生並改善神經元連接性[62]。BDNF也是蛋白synapsin-1的媒介,它通過加速神經元纖維的發育和維持突觸的接觸來改善突觸的形成[63]。Meng團隊[64]進行的一項研究中,在780 nm的光照射下,胚胎大鼠的神經組織中觀察到了更緊密的分支和纖維之間的互連性,表明這些蛋白質的活性增強。BDNF也與神經塑性適應的改善有關,這在腦外傷和中風的情況下尤其重要[65]。
如果可以最終證明LLLT刺激人類以及小鼠的神經可塑性和突觸形成,那麼就已經為廣泛人體臨床應用打開了大門[66]。神經可塑性受損或異常可能與多種腦部疾病有關,例如阿滋海默症、精神疾病、中風、頭部外傷和成癮[67-71]。
結論
治療中風或頭部外傷所致腦損傷的藥物試驗在很大程度上不算成功。儘管學術實驗室和業界投入了大量資金和研究,但阻止阿茲海默症和帕金森氏病的藥物進展仍不明顯。儘管諸如抗抑憂鬱症之類的精神科藥物已經很成熟,並且是世界上最暢銷的藥物之一,但它們的有效性令人失望,並且有很高的機率引發令人痛苦的副作用。考慮到LLLT的既定安全性、無明顯的不良作用以及相對的成本效益,現在該是正視LLLT對大腦疾病療效的時候了。
總體而言,LLLT對治療大腦疾病的前景非常樂觀,相關的治療機轉也被大量研究證實。在動物模型研究中獲得了非常正面的結果,並且到目前為止已經進行了許多小型人體臨床試驗,因此,有必要對LLLT及其在大腦神經系統疾病中的應用進行更廣泛的臨床測試,而使其成為一種普及使用的治療方法。
衍伸閱讀:紅光低能量光療法(LLLT)對健康的驚人好處/陳風河博士
參考資料
[1] L.F. De Freitas, M.R. Hamblin, Proposed mechanisms of photobiomodulation or low-level light therapy, IEEE J. Sel. Top. Quantum Electron. 22 (2016) 7000417.
[2] N. Lane, Cell biology: power games, Nature 443 (2006) 901–903.
[3] Mintzopoulos D, Gillis TE, Tedford CE, Kaufman MJ, Effects of Near-Infrared Light on Cerebral Bioenergetics Measured with Phosphorus Magnetic Resonance Spectroscopy. Photomed. Laser Surg 2017, 35, 395–400.
[4] Ando T et al., Comparison of Therapeutic Effects between Pulsed and Continuous Wave 810-nm Wavelength Laser Irradiation for Traumatic Brain Injury in Mice. PLoS ONE 2011, 6, e26212–e26220.
[5] Salehpour F et al., Transcranial low-level laser therapy improves brain mitochondrial function and cognitive impairment in D-galactose–induced aging mice. Neurobiol. Aging 2017, 58, 140–150.
[6] Kann O, The interneuron energy hypothesis: Implications for brain disease. Neurobiol. Dis 2016, 90, 75–85.
[7] Cao X et al., Astrocyte-derived ATP modulates depressive-like behaviors. Nat. Med 2013, 19, 773–777.
[8] Lyons DN et al., A Mild Traumatic Brain Injury in Mice Produces Lasting Deficits in Brain Metabolism. J. Neurotrauma 2018, 35, 2435–2447.
[9] Briston T; Hicks AR, Mitochondrial dysfunction and neurodegenerative proteinopathies: Mechanisms and prospects for therapeutic intervention. Biochem. Soc. Trans 2018, 46, 829–842.
[10] Swerdlow RH, Burns JM, Khan SM, The Alzheimer’s disease mitochondrial cascade hypothesis: Progress and perspectives. Biochim. Biophys. Acta 2014, 1842, 1219–1231.
[11] G.B. Waypa, K.A. Smith, P.T. Schumacker, O2 sensing, mitochondria and ROS signaling: the fog is lifting, Mol. Asp. Med. 47-48 (2016) 76–89.
[12] Y. Zhao, P.M. Vanhoutte, S.W. Leung, Vascular nitric oxide: beyond eNOS, J. Pharmacol. Sci. 129 (2015) 83–94.
[13]Wang X et al., Interplay between up-regulation of cytochrome-coxidase and hemoglobin oxygenation induced by near-infrared laser. Sci. Rep 2016, 6, 30540.
[14]Lee HI et al., Pretreatment with light-emitting diode therapy reduces ischemic brain injury in mice through endothelial nitric oxide synthase-dependent mechanisms. Biochem. Biophys. Res. Commun 2017, 486, 945–950.
[15]Charriaut-Marlangue C et al., Nitric oxide signaling in the brain: A new target for inhaled nitric oxide? Ann. Neurol 2013, 73, 442–448.
[16]Kisler K, Nelson AR, Montagne A, Zlokovic BV, Cerebral blood flow regulation and neurovascular dysfunction in Alzheimer disease. Nat. Rev. Neurosci 2017, 18, 419–434.
[17] M.D. Skopin, S.C. Molitor, Effects of near-infrared laser exposure in a cellular model of wound healing, Photodermatol. Photoimmunol. Photomed. 25 (2009) 75–80.
[18] D.W. Barrett, F. Gonzalez-Lima, Transcranial infrared laser stimulation produces beneficial cognitive and emotional effects in humans, Neuroscience 230 (2013) 13–23.
[19] G. Dougal, S.Y. Lee, Evaluation of the efficacy of low-level light therapy using 1072 nm infrared light for the treatment of herpes simplex labialis, Clin. Exp. Dermatol. 38 (2013) 713–718.
[20] F. Vatansever, M.R. Hamblin, Far infrared radiation (FIR): its biological effects and medical applications, Photonics Lasers Med. 4 (2012) 255–266.
[21] E. Palazzo, F. Rossi, V. de Novellis, S. Maione, Endogenous modulators of TRP channels, Curr. Top. Med. Chem. 13 (2013) 398–407.
[22] R. Planells-Cases et al., Complex regulation of TRPV1 and related thermo-TRPs: implications for therapeutic intervention, Adv. Exp. Med. Biol. 704 (2011) 491–515.
[23] M.J. Caterina, Transient receptor potential ion channels as participants in thermosensation and thermoregulation, Am. J. Physiol. Regul. Integr. Comp. Physiol. 292 (2007) R64–R76.
[24] P. Cassano, S.R. Petrie, M.R. Hamblin, T.A. Henderson, D.V. Iosifescu, Review of transcranial photobiomodulation for major depressive disorder: targeting brain metabolism, inflammation, oxidative stress, and neurogenesis, Neurophotonics 3 (2016) 031404.
[25] F. Tian, S.N. Hase, F. Gonzalez-Lima, H. Liu, Transcranial laser stimulation improves human cerebral oxygenation, Lasers Surg. Med. 48 (2016) 343–349.
[26] T. Ando, W. Xuan, T. Xu, T. Dai, S.K. Sharma, G.B. Kharkwal, Y.Y. Huang, Q. Wu, M.J. Whalen, S. Sato, M. Obara, M.R. Hamblin, Comparison of therapeutic effects between pulsed and continuous wave 810-nm wavelength laser irradiation for traumatic brain injury in mice, PLoS One 6 (2011), e26212.
[27] Zhang L; Zhang Y; Xing D LPLI inhibits apoptosis upstream of Bax translocation via a GSK-3betainactivation mechanism. J. Cell. Physiol 2010, 224, 218–228.
[28] Ling Q; Meng C; Chen Q; Xing D Activated ERK/FOXM1 pathway by low-power laser irradiation inhibits UVB-induced senescence through down-regulating p21 expression. J. Cell. Physiol 2014, 229, 108–116.
[29] Eells JT; Gopalakrishnan S; Valter K Near-Infrared Photobiomodulation in Retinal Injury and Disease. Adv. Exp. Med. Biol 2016, 854, 437–441.
[30] Eells JT; Henry MM; Summerfelt P; Wong-Riley MTT; Buchmann EV; Kane M; Whelan NT; Whelan HT Therapeutic photobiomodulation for methanol-induced retinal toxicity. Proc. Natl. Acad. Sci. USA2003, 100, 3439–3444.
[31] Huang YY; Nagata K; Tedford CE; McCarthy T; Hamblin MR Low-level laser therapy (LLLT) reduces oxidative stress in primary cortical neurons in vitro. J. Biophotonics 2013, 6, 829–838.
[32] Sharma SK; Kharkwal GB; Sajo M; Huang Y-Y; De Taboada L; McCarthy T; Hamblin MR Dose Response Effects of 810 nm Laser Light on Mouse Primary Cortical Neurons. Lasers Surg. Med 2011, 43, 851–859.
[33] Huang YY; Nagata K; Tedford CE; Hamblin MR Low-level laser therapy (810 nm) protects primary cortical neurons against excitotoxicity in vitro. J. Biophotonics 2014, 7, 656–664.
[34] Roomruangwong C et al., A neuroimmune, neuro-oxidative and neuro-nitrosative model of prenatal and postpartum depression. Prog. Neuro-Psychopharmacol. Boil. Psychiatry 2018, 81, 262–274.
[35] Rodríguez-Rodríguez A et al., Oxidative Stress in Traumatic Brain Injury. Curr. Med. Chem 2014, 21, 1201–1211.
[36] Wu J et al., The Role of Oxidative Stress and Inflammation in Cardiovascular Aging. BioMed Res. Int 2014, 2014, 615312.
[37] Wang X et al., Oxidative stress and mitochondrial dysfunction in Alzheimer’s disease. Biochim. Biophys. Acta 2014, 1842, 1240–1247.
[38] Steinhubl SR Why Have Antioxidants Failed in Clinical Trials? Am. J. Cardiol 2008, 101, S14–S19.
[39] Rahal A et al., Oxidative Stress, Prooxidants, and Antioxidants: The Interplay. BioMed Res. Int 2014, 2014, 761264.
[40] Ristow M et al., Antioxidants prevent health-promoting effects of physical exercise in humans. Proc. Natl. Acad. Sci. USA 2009, 106, 8665–8670.
[41] Salehpour F; Farajdokht F; Erfani M; Sadigh-Eteghad S; Shotorbani SS; Karimi P; Rasta SH; Mahmoudi J; Hamblin MR Transcranial near-infrared photobiomodulation attenuates memory impairment and hippocampal oxidative stress in sleep-deprived mice. Brain Res 2018, 1682, 36–43.
[42] Bartos A et al., Pre-conditioning with near infrared photobiomodulation reduces inflammatory cytokines and markers of oxidative stress in cochlear hair cells. J. Biophotonics 2016, 9, 1125–1135.
[43] Mungrue IN; Husain M; Stewart DJ The Role of NOS in Heart Failure: Lessons from Murine Genetic Models. Heart Fail. Rev 2002, 7, 407–422.
[44] Assis L; Moretti AI; Abrahao TB; de Souza HP; Hamblin MR; Parizotto NA Low-level laser therapy (808 nm) contributes to muscle regeneration and prevents fibrosis in rat tibialis anterior muscle after cryolesion. Lasers Med. Sci 2013, 28, 947–955.
[45] Cury V et al., Low level laser therapy increases angiogenesis in a model of ischemic skin flap in rats mediated by VEGF, HIF-1alpha and MMP-2. J. Photochem. Photobiol. B Boil 2013, 125, 164–170.
[46] Walski T; Drohomirecka A; Bujok J; Czerski A; Wąż G; Trochanowska-Pauk N; Gorczykowski M; Cichon R; Komorowska M Low-Level Light Therapy Protects Red Blood Cells Against Oxidative Stress and Hemolysis during Extracorporeal Circulation. Front. Physiol 2018, 9, 647.
[47] Sarandol A; Sarandol E; Eker SS; Erdinc S; Vatansever E; Kirli S Major depressive disorder is accompanied with oxidative stress: Short-term antidepressant treatment does not alter oxidative–antioxidative systems. Hum. Psychopharmacol. Clin. Exp 2007, 22, 67–73.
[48] Chen Z; Zhong C Oxidative stress in Alzheimer’s disease. Neurosci. Bull 2014, 30, 271–281.
[49] Lim W et al., Modulation of lipopolysaccharide-induced NF-kappaB signaling pathway by 635 nm irradiation via heat shock protein 27 in human gingival fibroblast cells. Photochem. Photobiol 2013, 89, 199–207.
[50] Yang Y; Gao L Celecoxib Alleviates Memory Deficits by Downregulation of COX-2 Expression and Upregulation of the BDNF-TrkB Signaling Pathway in a Diabetic Rat Model. J. Mol. Neurosci 2017, 62, 188–198.
[51] Chen AC-H et al., Low-Level Laser Therapy Activates NF-kB via Generation of Reactive Oxygen Species in Mouse Embryonic Fibroblasts. PLoS ONE 2011, 6, e22453.
[52] Chen AC-H et al., Effects of 810-nm Laser on Murine Bone-Marrow-Derived Dendritic Cells. Photomed. Laser Surg 2011, 29, 383–389.
[53] Yamaura M et al., Low level light effects on inflammatory cytokine production by rheumatoid arthritis synoviocytes. Lasers Surg. Med 2009, 41, 282–290.
[54] Filiano AJ; Gadani SP; Kipnis J Interactions of innate and adaptive immunity in brain development and function. Brain Res 2015, 1617, 18–27.
[55] Orihuela R; McPherson CA; Harry GJ Microglial M1/M2 polarization and metabolic states. Br. J. Pharmacol 2016, 173, 649–665.
[56] Fernandes KPS et al, Nunes FD Photobiomodulation with 660-nm and 780-nm laser on activated J774 macrophage-like cells: Effect on M1 inflammatory markers. J. Photochem. Photobiol. B Boil 2015, 153, 344–351.
[57] Singer AC et al., Noninvasive 40-Hz light flicker to recruit microglia and reduce amyloid beta load. Nat. Protoc2018, 13, 1850–1868.
[58] Bergmann O et al., Adult Neurogenesis in Humans. Cold Spring Harb. Perspect. Biol2015, 7, a018994.
[59] Oron A et al., Low-Level Laser Therapy Applied Transcranially to Rats after Induction of Stroke Significantly Reduces Long-Term Neurological Deficits. Stroke 2006, 37, 2620–2624.
[60] Xuan W et al., Transcranial low-level laser therapy enhances learning, memory, and neuroprogenitor cells after traumatic brain injury in mice. J. Biomed. Opt 2014, 19, 108003.
[61] Yang J; Harte-Hargrove LC; Siao C-J; Marinic T; Clarke R; Ma Q; Jing D; Lafrancois JJ; Bath KG; Mark W; et al. ProBDNF negatively regulates neuronal remodeling, synaptic transmission and synaptic plasticity in hippocampus. Cell Rep 2014, 7, 796–806.
[62] Yan X; Liu J; Zhang Z; Li W; Sun S; Zhao J; Dong X; Qian J; Sun H Low-level laser irradiation modulates brain-derived neurotrophic factor mRNA transcription through calcium-dependent activation of the ERK/CREB pathway. Lasers Med. Sci 2017, 32, 169–180.
[63] Barbieri R; Contestabile A; Ciardo MG; Forte N; Marte A; Baldelli P; Benfenati F; Onofri F Synapsin I and Synapsin II regulate neurogenesis in the dentate gyrus of adult mice. Oncotarget 2018, 9, 18760–18774.
[64] Meng C; He Z; Xing D Low-Level Laser Therapy Rescues Dendrite Atrophy via Upregulating BDNF Expression: Implications for Alzheimer’s Disease. J. Neurosci 2013, 33, 13505–13517.
[65] Wang Y; Huang Y-Y; Wang Y; Lyu P; Hamblin MR Photobiomodulation of human adipose-derived stem cells using 810 nm and 980 nm lasers operates via different mechanisms of action. Biochim. Biophys. Acta 2016, 1861, 441–449.
[66] Forrest MP; Parnell E; Penzes P Dendritic structural plasticity and neuropsychiatric disease. Nat. Rev. Neurosci 2018, 19, 215–234.
[67] Nahum M; Lee H; Merzenich MM Principles of Neuroplasticity-Based Rehabilitation. Pathol. Situat. Dis 2013, 207, 141–171.
[68] Kuhn M; Weidenauer A; Pezawas L Neuroplasticity and memory formation in major depressive disorder: An imaging genetics perspective on serotonin and BDNF. Restor. Neurol. Neurosci 2014, 32, 25–49.
[69] Felling RJ; Song H Epigenetic mechanisms of neuroplasticity and the implications for stroke recovery. Exp. Neurol 2015, 268, 37–45.
[70] Tomaszczyk JC; Green NL; Frasca D; Colella B; Turner GR; Christensen BK; Green RE A. Negative Neuroplasticity in Chronic Traumatic Brain Injury and Implications for Neurorehabilitation. Neuropsychol. Rev 2014, 24, 409–427.
[71] Creed M; Pascoli VJ; Luscher C Refining deep brain stimulation to emulate optogenetic treatment of synaptic pathology. Science 2015, 347, 659–664.